

- L’analyse génétique est symbolisée par l’étude de la variation d’un individu à l’autre d’une répétition en tandem d’unités de bases appelées également les marqueurs, marqueurs STRs ou microsatellites. Il faut donc que le prélèvement du matériel biologique sous forme de trace indiciale ou corporelle subisse toute une série de traitements biologiques ou chimiques afin de permettre le processus identificatoire.
- La première étape est la purification de l’échantillon, soit l’extraction et la séparation de l’ADN des globules rouges et autres impuretés[1].
- Après la phase d’extraction, deux possibilités existent. La quantification de l’ADN est suffisante et l’analyse proprement dite peut débuter, ou elle est insuffisante impliquant l’usage de la technique PCR. La PCR – Polymerase Chain Reaction – est une technique d’amplification de l’ADN, soit de copiage moléculaire, qui permet d’outrepasser les limites quantitatives de l’ADN[2]. Depuis l’année 2000, la technique PCR est systématiquement utilisée par les Instituts de médecine légale suisses jusqu’à une duplication de 36 fois.
- La phase d’extraction de l’ADN ou la technique de PCR est suivie d’une électrophorèse. Aujourd’hui, les laboratoires de médecine légale utilisent le séquenceur automatique permettant d’effectuer une électrophorèse automatisée et d’obtenir un résultat informatisé. Le séquenceur automatique fourni sous forme de chiffres deux marqueurs – l’un correspond à l’allèle transmis par le père et l’autre par la mère – et ceci pour chaque séquence répétitive. A noter que plus le temps s’écoule avant d’obtenir le résultat pour un marqueur, plus l’allèle est long et donc, plus le nombre de répétitions est important avec un meilleur taux de discrimination[3].
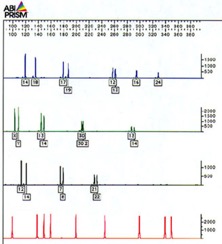
Cette image représente le résultat d’une chromatographie. Les pics représentent les marqueurs, il peut y avoir un ou deux caractères par marqueur, soit un ou deux nombres, selon que l’allèle hérité du père et celui de la mère sont identiques (locus homozygote) ou différents (locus hétérozygote). Chaque ligne de l’électrophorégramme correspond à une couleur de fluochrome.
- Outre le gain de temps, le séquenceur automatique a l’avantage de pouvoir lire jusqu’à mille nucléotides alors que la méthode manuelle d’électrophorèse se limite à 300 nucléotides[4].
- En fonction de l’emplacement et de la taille des microsatellites mis en évidence par le séquenceur automatique, il est possible de réaliser un profil génétique[5].
[1] A ce sujet: Altendorfer, p. 35; Coquoz, Comte, Hall, Hicks, Taroni, p. 42-43; Klumpe, p. 16-17. [2] Altendorfer, p. 42-43; BSK-StPO-Fricker, Maeder, art. 255 ss N 13; Busch, p. 637; Coquoz, Comte, Hall, Hicks, Taroni, p. 45-46; Klumpe, p. 36-37; Lavergne, p. 27; Ruckstuhl, Dittmann, Arnold, p. 544. [3] Coquoz, Comte, Hall, Hicks, Taroni, p. 40-51. [4] Ameziane, Bogard, Lamoril, p. 409-410. [5] Buquet, p. 178; Coquoz, Comte, Hall, Hicks, Taroni, p. 40-41; Kaye, p. 679; Lavergne, p. 28.